

The ground beneath our feet may seem stable, but in many regions, it is constantly shifting. As climate change speeds up and cities expand into hilly and mountainous areas, landslides are happening more often and with greater force. This growing threat calls for smarter and more reliable tools to understand natural hazards. For years, most risk models have focused on surface factors like slope angle and rainfall. While useful, these models often overlook what lies deeper below the surface—the geological “DNA” of the land. As a result, their predictions can fall short. This is where the geological complexity index (GCI) makes a real difference. Instead of relying on a few basic variables, GCI uses a multi-parameter approach. It considers tectonic history, variations in rock types, and the strength of underground structures. By connecting these deeper factors, the geological complexity index is reshaping landslide risk modeling. It is also helping scientists predict hazards with greater confidence and clarity. Why Geological Complexity Index Matters Now We are now living in the age of cascading disasters. One heavy rainfall no longer causes just a flood. In high-altitude or geologically active regions, it often sets off a chain reaction. Slopes collapse. Land shifts. Entire landscapes fail in sequence. For decades, scientists focused mainly on the trigger. Rainfall. Earthquakes. Extreme weather. Meanwhile, the ground beneath was treated as something stable and uniform. But thanks to modern remote sensing and GIS modeling, we now know that assumption was wrong. The Earth is not uniform at all. It is layered, fractured, and constantly evolving. This is where the Geological Complexity Index comes in. It offers a way to measure the hidden “inner chaos” of the terrain. Yes, a steep slope is risky. But a steep slope made of fractured limestone, resting on an active fault line, is far more dangerous. GCI captures this difference. It assigns a numerical value to geological complexity, adding a deeper and more realistic dimension to landslide risk assessment. And this matters now more than ever. Traditional models are reaching their limits. Adding more rainfall data alone no longer improves predictions significantly. To truly boost accuracy, we need to understand how complex the ground itself is. GCI is not just another tool for terrain research. It is becoming a life-saving foundation for the next generation of early warning systems. What Is the Geological Complexity Index? At its core, the geological complexity index measures how unstable a geological environment really is. It does not rely on a single data point. Instead, it brings together several high-impact factors into one clear metric. In other words, it looks at the big picture. It helps explain how a landscape is likely to behave when pressure builds. As stress increases, this combined view makes it easier to understand why some areas fail while others remain stable. The Four Pillars of GCI To understand GCI, we have to break it down into its constituent components. Think of these as the “ingredients” that define the volatility of the terrain: How GCI is Calculated Calculating the GCI is a feat of multi-parameter modeling. It typically utilizes the Analytic Hierarchy Process. In simple terms, AHP is a structured technique for organizing and analyzing complex decisions. Researchers take data from Digital Elevation Models, satellite imagery, and field surveys, then assign “weights” to each pillar. For example, in a region known for massive earthquakes, the seismicity component might be weighted more heavily than lithologic diversity. By combining these weighted factors, we get a single, powerful index that represents the total geological “stress” of the area. How GCI Improves Landslide Risk Modeling The evidence is clear in the data. Recent breakthroughs back this up. One standout example is Zhang’s study on landslide susceptibility in mountainous reservoir regions. The research shows that adding the geological complexity index greatly improves prediction accuracy. In fact, it outperforms traditional models by a noticeable margin. Case Evidence: The Zhang Study Effect In many traditional landslide risk modeling scenarios, the “Area Under the Curve” (AUC)—a standard metric for model success—often hovers around 0.7 to 0.8. However, when Zhang and his team integrated GCI into their hazard evaluation, the predictive power often jumped into the 0.85–0.92 range. Why the massive jump? Because traditional models often struggle with “false positives” (predicting a slide where none occurs) or, more dangerously, “false negatives.” GCI refines the model by explaining why two identical-looking slopes behave differently. One slope might have a GCI of 0.3 (stable), while another—due to hidden faulting—might have a GCI of 0.8. GCI vs. Traditional Machine Learning Models We are now seeing a surge in machine learning applications in geology. While ML is great at finding patterns, it is only as good as the features you feed it. Terrain and Geological Interactions The relationship between terrain complexity research and GCI is a symbiotic one. You can’t have a deep understanding of one without the other. The Influence of Terrain Features When we look at a Digital Elevation Model, we see several key features: Integrating GCI with Terrain Complexity In modern landslide susceptibility indexing, we look at the interaction between the surface (terrain) and the subsurface (geology). For example, a “planar” terrain might seem safe, but if the GCI indicates high fault density underneath, that surface is essentially sitting on a cracked foundation. By integrating these, we move from 2D maps to 3D “volumetric” risk assessments. This is the frontier of natural hazard analysis: seeing the mountain not just as a shape, but as a living, complex mechanical system. Beyond GCI — Integrating Climate & Human Triggers As powerful as the geological complexity index is, it doesn’t exist in a vacuum. It represents the “pre-disposition” of the land, but it still needs a “trigger.” The Synergy of Rainfall and GCI Rainfall is the most common trigger for landslides. However, a high-GCI area is significantly more sensitive to rainfall. In a low-complexity zone, it takes 200mm of rain to trigger a slide. In a high-complexity zone, 50mm might be enough to lubricate existing faults

Cardiovascular System Explained: How Our Heart & Blood Keep Us Alive
Think of a city that never shuts down. It’s always active, always moving. To stay alive, it needs nonstop deliveries of food, water, and oxygen. At the same time, waste has to be cleared fast and efficiently. Inside our body, that city is made up of organs and tissues. The system that keeps everything running is the cardiovascular system. This system is a powerful, nonstop network of the heart, blood, and blood vessels. It works day and night without breaks. Every second, it delivers oxygen and nutrients to our cells, from our brain down to our toes. It also carries away waste before it can cause harm. Without this system, the body would shut down within minutes. In this guide, we’ll take a clear and simple journey through this inner highway. First, we’ll look at the structure of the heart and how it works. Then, we’ll explore the different types of blood vessels and their roles. After that, we’ll break down the cardiac cycle and its steady rhythm. Finally, we’ll see how our body controls blood pressure to keep us active, balanced, and full of energy. What is the cardiovascular system? The cardiovascular system, often called the circulatory system, works like the body’s main delivery network. Its core job is simple but powerful: move fluids through the body in a continuous, closed loop under pressure. At first, it may sound like it’s all about the heart. However, the system only works because three key parts operate together. Think of it as a well-coordinated team. First, there is the pump. The heart generates the force that keeps everything moving. Next comes the fluid. Blood carries oxygen, nutrients, hormones, and waste from one place to another. Finally, there are the pipes. Blood vessels—arteries, veins, and capillaries—form the pathways that guide blood throughout the body. Together, these three components keep life flowing, second by second. Why It Is Essential for Life The cardiovascular system serves as the body’s ultimate multitasker. Its primary role is homeostasis—the maintenance of a stable internal environment. It achieves this through: Blood: The Life-Sustaining Fluid of the Cardiovascular System What Makes Blood So Special? Blood is far more than just a red liquid flowing through our veins. It’s a living tissue composed of cells suspended in a liquid called plasma. If you were to separate blood in a test tube, you’d see it naturally divides into layers, revealing its hidden complexity. Composition: Cells + Plasma Plasma makes up about 55% of our blood volume. This yellowish fluid is mostly water (around 90%), but it’s packed with dissolved proteins, glucose, hormones, electrolytes, and clotting factors. Think of plasma as the delivery truck that carries everything our cells need. The remaining 45% consists of cellular components: Functions: Transport, Immunity, and Regulation Blood performs three critical jobs simultaneously: Heart Anatomy—The Central Pump Our heart sits in our chest cavity, slightly left of center, nestled between our lungs. About the size of our fist, it beats approximately 100,000 times daily, pumping blood with remarkable efficiency. Three protective layers shield this vital organ: The pericardium is the tough outer sac that anchors our heart and prevents overfilling. It’s like a protective sleeping bag that holds our heart in place while allowing movement. The myocardium is the thick muscular middle layer that does the actual pumping. These specialized cardiac muscle cells contract in perfect coordination, generating the force needed to push blood throughout our entire body. The endocardium is the smooth inner lining that prevents blood from sticking to the heart walls. This slick surface ensures blood flows smoothly without clotting. Chambers and Valves: The Four-Room Design Our heart functions as two pumps working side by side, divided into four chambers: The right atrium receives deoxygenated blood returning from our body through two large veins called the superior and inferior vena cavae. This thin-walled chamber acts as a receiving room. The right ventricle pumps this oxygen-poor blood to our lungs through the pulmonary arteries. It doesn’t need to generate as much pressure since the lungs are nearby. The left atrium receives freshly oxygenated blood from our lungs via four pulmonary veins. Again, it’s a temporary holding chamber. The left ventricle is the powerhouse. Its thick, muscular walls generate enough force to send blood on a journey through our entire body. This chamber works the hardest and is prone to problems when blood pressure stays elevated. Four valves act as one-way doors, ensuring blood flows in the right direction: When these valves malfunction, you might hear a heart murmur—the sound of blood flowing backward or through a narrowed opening. Major Vessels: The Highway Connections The aorta is our body’s main artery, emerging from the left ventricle. This massive vessel branches into smaller arteries that reach every corner of our body, from our brain to our toes. The vena cavae are the two largest veins returning blood to the heart. The superior vena cava drains our upper body, while the inferior vena cava handles everything below our heart. The pulmonary arteries carry deoxygenated blood from our right ventricle to both lungs. Interestingly, these are the only arteries in our body that carry oxygen-poor blood. The pulmonary veins return oxygenated blood from our lungs to the left atrium, completing the pulmonary circuit. Blood Vessels and Circuits Arteries vs. Veins vs. Capillaries Our circulatory system anatomy includes three types of blood vessels, each designed for specific tasks: Arteries are thick-walled, muscular vessels that carry blood away from our heart. They must withstand high pressure, so their walls contain elastic fibers and smooth muscle. You can feel an artery’s pulse because it expands slightly with each heartbeat. The largest arteries branch into smaller arterioles that control blood flow to different organs. Veins return blood to our heart under much lower pressure. Their walls are thinner and less elastic than arteries. Many veins, especially in our legs, contain one-way valves that prevent backward flow when we’re standing. When these valves weaken, we develop varicose veins. Capillaries are microscopic vessels where
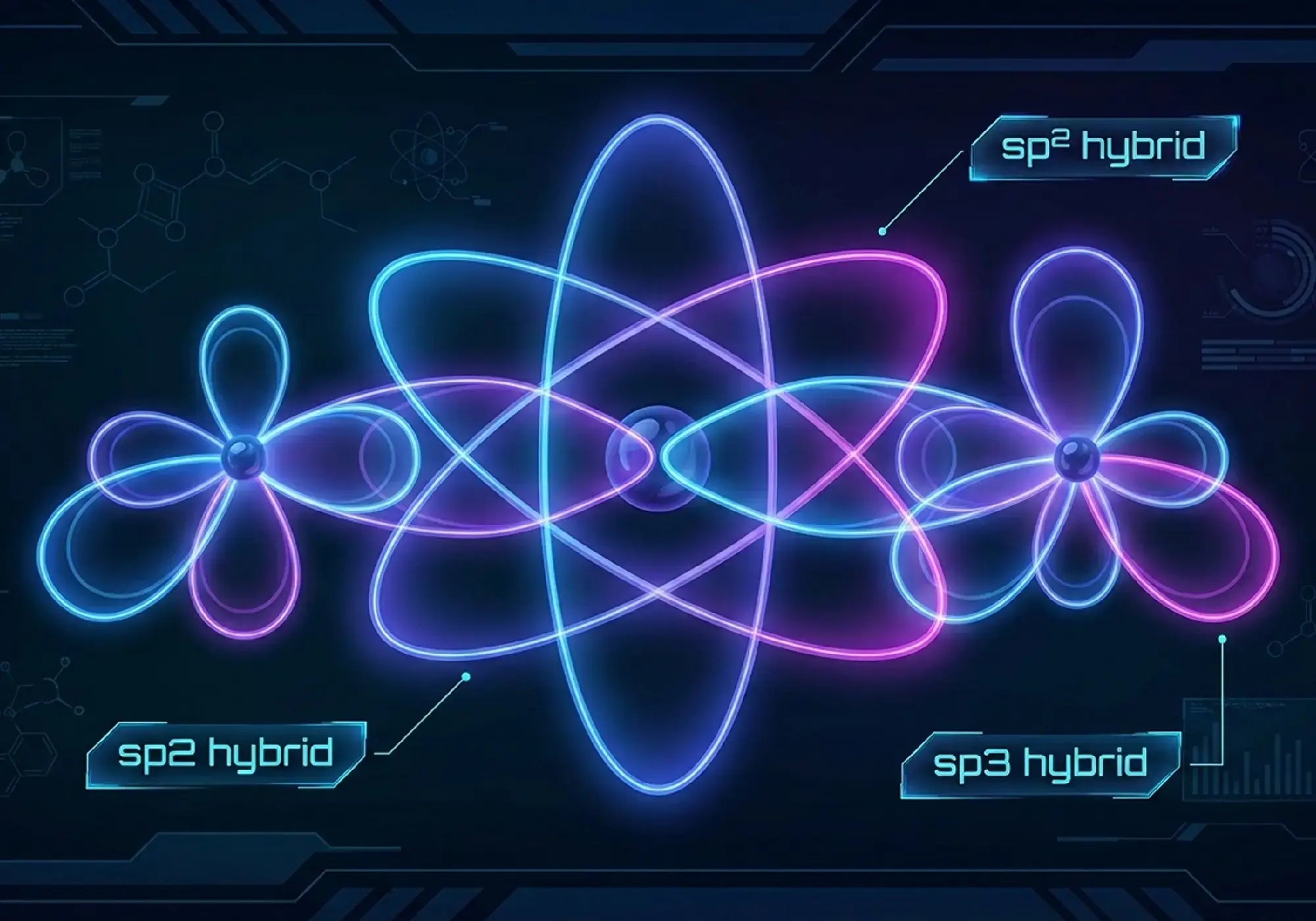
The Secret Life of Organic Chemistry Bonding: Unlocking Hybridization, Shapes & Electron Clouds
Curious about why carbon can form millions of compounds while most of the other elements barely make a few? The secret lies in organic chemistry bonding the invisible framework that builds every molecule around you, from the DNA in our cells to the caffeine in our morning coffee. Understanding atoms is not just about memorizing formulas. It’s about seeing electron clouds, feeling how orbitals overlap, and visualizing why molecules twist, bend, and react. Hybridization and molecular geometry are not just abstract ideas; they explain why diamonds are hard, gasoline burns, and our phone screen responds to our touch. In this deep dive, we will explore sigma and pi bonds, decode hybridization from sp³ to sp, and uncover how electron density maps reveal the real nature of chemical bonding. Whether you are aiming to ace organic chemistry or are simply curious, this guide will change the way you see molecules forever. Covalent Bonds: A Quick Refresher Covalent bonds happen when atoms share electrons to become stable. But here’s the twist textbooks often miss: electrons don’t just sit between atoms like tiny planets. The real magic is orbital overlap, when atomic orbitals merge to form a shared space for electrons. Imagine two soap bubbles touching. The area where they overlap? That’s your bond. The bigger the overlap, the stronger the connection. This shared space packs electron density right between the nuclei, holding them together with electrostatic attraction. Sigma (σ) vs. Pi (π) Bonds There are two fundamental types of covalent bonds, and understanding their difference is crucial for mastering organic chemistry bonding: Sigma (σ) bonds form through head-on orbital overlap. Picture two orbitals meeting face-to-face along the axis connecting the atoms. This creates a cylindrical cloud of electron density directly between the nuclei. Every single bond you see is a sigma bond. Pi (π) bonds form through side-by-side overlap of p orbitals. Imagine two parallel orbitals touching along their sides, creating electron density above and below the bonding axis. Pi bonds always come as additions to sigma bonds in double and triple bonds. Here’s the visual breakdown: The sigma framework acts as the skeleton of organic molecules, while pi clouds add reactivity and rigidity. Why Sigma Bonds Allow Rotation, Pi Don’t This is where it gets fascinating. Sigma bonds allow free rotation because their electron cloud is evenly spread around the bond axis. Twist a single bond all the way around, and the orbital overlap stays the same; nothing snaps. Pi bonds, on the other hand, are stuck. Their side-by-side p orbital overlap would break if the bond rotated, destroying the connection. That’s why double bonds make molecules rigid and give rise to cis-trans isomers. Think of the pi cloud like a molecular handcuff, locking the shape in place. Recognizing how bonds rotate is key to predicting how molecules twist, fold, and react in organic chemistry. Hybridization Theory: From Orbitals to Shapes In the 1930s, Linus Pauling cracked a big mystery: if carbon has two types of atomic orbitals—one s and three p—why do its four bonds in methane all look the same? His answer was hybridization. Before bonding, atomic orbitals combine mathematically to form new, equivalent hybrid orbitals. This isn’t a physical mix; it’s a quantum idea that predicts molecular shapes and bonding patterns more accurately. Valence bond theory uses hybridization to explain molecule shapes, while molecular orbital theory shows the full electronic structure. Both are correct, but hybridization is easier to grasp, especially when studying organic chemistry bonding. sp, sp², sp³ Explained with Shapes & Angles To understand hybridization explained for the organic world, we look at how many “electron groups” (atoms or lone pairs) surround an atom. sp³ Hybridization: One s orbital + three p orbitals = four equivalent sp³ orbitals arranged tetrahedrally. Bond angle: 109.5°. This is your standard carbon in alkanes, with maximum electron repulsion balance according to VSEPR theory. sp² Hybridization: One s orbital + two p orbitals = three sp² orbitals in a trigonal planar arrangement. Bond angle: 120°. One p orbital remains unhybridized and perpendicular to the plane, ready to form pi bonds. This is the carbon in alkenes. sp Hybridization: One s orbital + one p orbital = two sp orbitals in a linear arrangement. Bond angle: 180°. Two p orbitals remain unhybridized for forming two pi bonds. This is the carbon in alkynes. The pattern? More s character means orbitals hold electrons closer to the nucleus, making atoms more electronegative and bonds stronger and shorter. Hybridization Types in Organic Chemistry Hybridization Orbitals Mixed Number of Hybrid Orbitals Molecular Shape Bond Angle Unhybridized p Orbitals Typical Organic Example sp³ 1 s + 3 p 4 sp³ Tetrahedral 109.5° 0 Alkanes (e.g., methane, ethane) sp² 1 s + 2 p 3 sp² Trigonal planar 120° 1 Alkenes (e.g., ethene) sp 1 s + 1 p 2 sp Linear 180° 2 Alkynes (e.g., ethyne) Lone Pairs and Deviations Hybridization isn’t just about bonding orbitals—lone pairs play a role too. They occupy hybrid orbitals and influence molecular shapes by repelling other electrons. Take ammonia (NH₃) as an example. Its nitrogen is sp³ hybridized with one lone pair. Normally, a tetrahedral angle would be 109.5°, but the lone pair pushes harder than bonding pairs. This compresses the H–N–H angle to 107°. Water (H₂O) takes it a step further. Oxygen is sp³ hybridized with two lone pairs. These lone pairs push strongly, squeezing the H–O–H angle to 104.5°. The molecule forms a bent shape, not a perfect tetrahedron, because lone pairs dominate the repulsion forces. This shows that hybridization predicts how orbitals arrange themselves. VSEPR theory fine-tunes the molecular geometry. It does so by accounting for electron-electron repulsion. Electron Clouds & Bonding Patterns in Organic Chemistry Bonding We often draw electrons as dots, but in reality, they exist as a “cloud” or electron density. 1. Electron Density Maps Electron density maps show where electrons actually hang out. Instead of rigid sticks, bonds appear as fuzzy clouds with areas of high and low concentration. When electron density is

Human Cell Atlas 2026: The Ultimate Guide to Comprehensive Cellular Mapping
Imagine having a full map of every cell in the human body, all 37 trillion of them. That is the bold idea behind the Human Cell Atlas 2026. It is a global project changing how we understand human biology at its deepest level. The Human Cell Atlas is not just another research study. Instead, it’s a massive collaboration. Thousands of scientists from six continents are working together to build the most detailed reference map of human cells ever created. Think of it like Google Maps for the human body. It shows where each cell lives, what it does, and its unique molecular fingerprint, from brain neurons to cells in the gut. The project began in 2016 as a big, almost unreal vision. Since then, it has grown into something very real. By 2026, researchers plan to finish the first complete draft of the atlas. This step could reshape how we detect diseases, design treatments, and personalize medicine. It’s not science fiction anymore. It is unfolding right now. Technologies Behind Cellular Mapping The Human Cell Atlas 2026 is only possible because of a powerful tech stack—one that honestly feels straight out of sci-fi. But to truly understand how scientists are mapping the human body, we need to zoom in on the tools making it happen. 1. Single-Cell Genomics Traditional biology studied cells in bulk, like analyzing an entire forest without distinguishing individual trees. Single-cell sequencing changed everything. This technology reads the genetic activity of individual cells, revealing hidden diversity we never knew existed. For example, what we once called “lung cells” are actually dozens of distinct types, each with unique functions. Some cells sense oxygen levels, others produce surfactant, and some coordinate immune responses. 2. Spatial Transcriptomics Knowing what a cell does is important, but knowing where it does it is equally crucial. Spatial transcriptomics maps gene activity while preserving the physical location of cells within tissues. This reveals how cells interact with neighbors and respond to their local environment. 3. Multi-Omics Integration The most powerful insights come from combining multiple data types—genomics, proteomics, metabolomics—into integrated multi-omics profiles. This holistic view captures cell identity and state with unprecedented precision, revealing how genetics, proteins, and metabolites work together. 4. Computational Tools & Data Portals Managing petabytes of cellular data requires sophisticated computational infrastructure. The HCA Data Portal serves as the central hub, offering standardized tools for analysis and visualization. Researchers can explore datasets, compare cell types across organs, and download raw data—all through user-friendly interfaces. How Cell Type Mapping Works Defining what makes a cell a type versus a state is not as simple as it seems. A liver cell, for example, is clearly a cell type. However, that same cell can shift through different states. It may be resting, dividing, or reacting to injury. In other words, the identity stays the same, but the behavior changes. To handle this complexity, the Human Cell Atlas relies on advanced ontologies and detailed annotation systems. These tools help scientists classify cells in a consistent way, even when the data comes from different studies. At the same time, data standardization remains a major hurdle. Labs around the world use different protocols, tools, and analysis methods. As a result, the same cell can look slightly different depending on where it is studied. To fix this, universal standards are essential. They ensure that a T cell identified in Boston is recognized as the same T cell mapped in Beijing. This consistency is what makes the atlas reliable, comparable, and truly global. Major Human Cell Atlas Projects and Key Achievements The HCA is not just a theory, it is delivering real maps. Organized by networks, it is churning out organ-specific gems. Organ-Specific Atlases The HCA tackles the body organ by organ. The Lung Biological Network mapped respiratory cells, revealing rare populations involved in COVID-19 susceptibility. The Gut Cell Atlas characterized intestinal cells, advancing understanding of inflammatory bowel disease. The Thymus Atlas illuminated immune cell development, while the Skeletal Atlas explored bone and cartilage biology. Each organ atlas is both a standalone resource and a piece of the larger human puzzle. First Draft Progress Toward HCA In 2023, the consortium announced significant progress toward the “first draft” of the complete atlas—covering major organs and tissues with sufficient depth to serve as initial reference maps. While far from complete, these drafts already enable disease biomarkers discovery and clinical applications. The 2026 General Meeting aims to consolidate these achievements, set priorities for HCA 2.0, and expand coverage to underrepresented tissues and populations. Insights Already Unlocked in Disease Research The atlas is not just future promise—it is delivering now. Researchers have identified new cell types involved in fibrosis, discovered cellular origins of cancer, mapped immune responses to infections, and found therapeutic targets for rare diseases. One striking example: comparing healthy and diseased tissue atlases revealed specific cell states linked to severe COVID-19, guiding treatment strategies and vaccine development. Ethical, Legal, and Social Dimensions of the Human Cell Atlas A map of the human body is a map of us. This brings up heavy questions. Sampling Diversity Early genomics efforts suffered from severe bias—most data came from people of European ancestry. The HCA actively addresses this by recruiting diverse donors across ethnicities, ages, and geographies. Global representation ensures the atlas reflects human diversity, not just a narrow slice. This matters clinically. Disease manifests differently across populations. Treatments developed from homogeneous data may not work universally. Privacy and Consent Collecting human tissue requires robust consent processes. The HCA follows strict ethical guidelines, ensuring donors understand how their samples will be used. Data is de-identified to protect privacy while remaining scientifically useful. Balancing ethical data sharing with individual privacy rights requires constant vigilance, especially as computational methods make re-identification increasingly possible. Global Equity in Science Who benefits from the atlas? Ideally, everyone. But there’s risk that high-income countries with advanced research infrastructure capture most benefits. The HCA works to build capacity in lower-resourced settings, providing training, tools, and collaborative opportunities. True equity means not